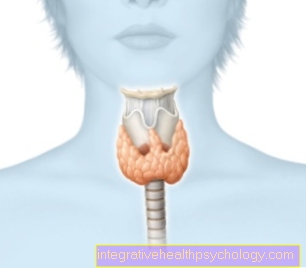
Tyrosinkinasen
Hva er en tyrosinkinase?
Tyrosinkinase er en spesifikk gruppe enzymer som er funksjonelt tilordnet proteinkinaser fra et biokjemisk synspunkt. Proteinkinaser overfører reversibelt (mulighet for omvendt reaksjon) fosfatgrupper til OH-gruppen (hydroksylgruppe) av aminosyren tyrosin. Fosfatgruppen overføres til hydroksylgruppen i tyrosinet til et annet protein.
Gjennom denne reversible fosforyleringen som er beskrevet, kan tyrosinkinaser avgjørende påvirke aktiviteten til proteiner og derfor spille en viktig rolle i signaltransduksjonsveier. Spesielt terapeutisk, for eksempel innen onkologi, brukes funksjonen av tyrosinkinaser som et mål for medikamenter.

Oppgaven og funksjonen
Tyrosinkinaser må først deles inn i membranbundne og ikke-membranbundne tyrosinkinaser for å forstå hvordan de fungerer.
Membranbundne tyrosinkinaser kan ha sin egen proteinkinaseaktivitet, idet kinasefunksjonen aktiveres som en del av reseptorkomplekset på cellemembranen.Ellers kan membranbundne tyrosinkinaser være funksjonelt knyttet til reseptorkomplekset, men kan ikke lokaliseres direkte i det. Tyrosinkinasen og reseptoren danner en binding gjennom hvilken et bestemt signal sendes videre til kinasen via reseptoren.
I tilfelle av en ikke-membranbundet tyrosinkinase, er dette enten i cytoplasmaet eller i en cellekjerne. Ulike eksempler på tyrosinkinaser kan navngis avhengig av den strukturelle utformingen med en tilhørende funksjon. Eksempler på membranbundne tyrosinkinaser er insulinreseptoren, EGF-reseptoren, NGF-reseptoren eller PDGF-reseptoren. Dette viser at signalet kaskader ved hjelp av tyrosinkinaser er viktige prosesser i menneskekroppen.
Insulinfrigivelsen fra bukspyttkjertelen i forbindelse med måltider reguleres via insulinreseptoren. EGF-reseptoren har spesifikke bindingssteder for flere ligander, blant dem er det verdt å nevne EGF eller TNF-alfa. Som proteinligand tar EGF (epidermal vekstfaktor) en enestående rolle som vekstfaktor (celleproliferasjon og differensiering). TNF-alpha er derimot en av de kraftigste inflammatoriske markørene i menneskekroppen og spiller en viktig diagnostisk rolle i diagnosen betennelse.
PDGF er i sin tur en vekstfaktor som frigjøres av trombocytter (blodplater), som induserer sårlukking og, ifølge dagens forskning, også bidrar til utviklingen av pulmonal hypertensjon.
Eksempler på ikke-membranbundne tyrosinkinaser er ABL1 og Janus-kinaser.
I prinsippet går en signalkaskade med viss informasjon alltid på samme stereotype måte når det gjelder tyrosinkinase. For det første må en passende ligand binde seg til en reseptor, som vanligvis ligger på overflaten av celler. Denne koblingen blir vanligvis etablert via en kongruent proteinstruktur av ligand og reseptor (lås og nøkkelprinsipp) eller via binding til visse kjemiske grupper i reseptoren (fosfat, sulfatgrupper, etc.). Koblingen endrer reseptorens proteinstruktur. Spesielt når det gjelder tyrosinkinaser, danner reseptoren homodimerer (to identiske proteinunderenheter) eller heterodimerer (to forskjellige proteinunderenheter). Denne såkalte dimeriseringen kan føre til en aktivering av tyrosinkinaser som, som allerede nevnt ovenfor, er lokalisert direkte i reseptoren eller på den cytoplasmiske siden (rettet mot innsiden av cellen) av reseptoren.
Aktivering knytter hydroksylgruppene av tyrosinrester i reseptoren med fosfatgrupper (fosforylering). Denne fosforyleringen skaper gjenkjennelsessteder for intracellulært lokaliserte proteiner som senere kan binde seg til dem. De gjør dette via spesifikke sekvenser (SH2-domener). Etter binding til fosfatgruppene utløses svært komplekse signalkaskader i cellekjernen, noe som igjen fører til fosforylering.
Det skal bemerkes at aktiviteten til proteiner kan påvirkes i begge retninger via fosforylering av tyrosinkinaser. På den ene siden kan disse aktiveres, men på den andre siden kan de også deaktiveres. Det kan sees at en ubalanse i tyrosinkinaseaktiviteten kan føre til en overstimulering av vekstfaktorassosierte prosesser, som til slutt tillater kroppens celler å formere seg og differensiere (tap av cellulært genetisk materiale). Dette er de klassiske prosessene for tumorutvikling.
Mangelfulle reguleringsmekanismer for tyrosinkinaser spiller også en avgjørende rolle i utviklingen av diabetes mellitus (insulinreseptor), arteriosklerose, pulmonal hypertensjon, visse former for leukemi (spesielt CML) eller ikke-småcellet lungekreft (NSCLC).
Finn ut alt om emnet her: Tumorsykdommer.
Hva er tyrosinkinasereseptoren?
Tyrosinkinase-reseptoren er en membranbasert reseptor, dvs. en reseptor forankret i cellemembranen Strukturelt sett er dette en reseptor med et transmembrankompleks. Dette betyr at reseptoren trekker gjennom hele cellemembranen og også har en ekstra- og intracellulær side.
Den spesifikke liganden binder seg til reseptoren på den ekstracellulære siden, alfa-underenheten, mens reseptorens katalytiske sentrum ligger på den intracellulære siden, β-underenheten. Det katalytiske sentrum representerer det aktive området av enzymet, der spesifikke reaksjoner finner sted.
Som allerede nevnt ovenfor, er strukturen til reseptoren vanligvis sammensatt av to proteinunderenheter (dimerer).
I tilfelle av insulinreseptoren, for eksempel, binder de to alfa-underenhetene insulinliganden. Etter ligandbinding er fosfatgrupper (såkalt fosforylering) bundet til spesifikke tyrosinrester (hydroksylgrupper). Dette genererte reseptorens tyrosinkinaseaktivitet. I det følgende kan ytterligere substratproteiner (f.eks. Enzymer eller cytokiner) inne i cellen aktiveres eller inaktiveres via fornyet fosforylering, og derved påvirke celleproliferasjon og differensiering.
Hva er en tyrosinkinasehemmer?
Såkalte tyrosinkinasehemmere (også: tyrosinkinasehemmere) er relativt nye medisiner som kan brukes til spesifikk behandling av mangelfull tyrosinkinase-aktivitet. De er klassifisert som kjemoterapeutiske midler og har sin opprinnelse på slutten av 1990-tallet og begynnelsen av 2000-tallet. De kan klassifiseres i forskjellige generasjoner og brukes til behandling av ondartede sykdommer.
Funksjonelt kan spesifikke prosesser forhindres av ubalanserte tyrosinkinaseaktiviteter. I prinsippet er fire forskjellige virkningsmekanismer mulig her. I tillegg til å konkurrere med ATP, er binding til reseptorens fosforyleringsenhet, til substratet eller allosterisk utenfor det aktive senteret også mulig. Effekten av tyrosinkinaseinhibitorene utløses ved binding til EGF-reseptoren og den påfølgende hemming av den enzymatiske aktiviteten til tyrosinkinasene.
Når det gjelder medisinsk historie, oppdaget oppdagelsen av den aktive ingrediensen imatinib som tyrosinkinasehemmere en enestående posisjon. Det brukes spesifikt i kronisk myeloid leukemi (CML), der det undertrykker tyrosinkinaseaktiviteten som patologisk skapes av en kromosomfusjon (Philadelphia-kromosom ved fusjon av kromosom 9 og 22).
Flere andre tyrosinkinasehemmere har blitt utviklet de siste årene. Den nåværende 2. generasjonen inneholder rundt ti tyrosinkinasehemmere.
Les mer om temaet her:
- Målrettet cellegift med tyrosinkinasehemmere
- Kronisk myeloid leukemi.
Til hvilke indikasjoner brukes de?
Tyrosinkinasehemmere brukes til forskjellige ondartede sykdommer. Imatinib brukes spesielt i kronisk myeloid leukemi. Andre mulige bruksområder er ikke-småcellet lungekreft (NSCLC), brystkreft og tykktarmskreft.
På grunn av den veldig selektive angrepsmekanismen til tyrosinkinaseinhibitorene tolereres de vanligvis bedre enn konvensjonelle kjemoterapeutiske midler. Likevel kan det også forventes bivirkninger her i detalj.
Finn ut mer om: Lungekreft.













.jpg)