Kromosomer
Definisjon - hva er kromosomer?
Den genetiske sammensetningen av en celle lagres i form av DNA (deoksyribonukleinsyre) og dens baser (adenin, tymin, guanin og cytosin). I alle eukaryote celler (dyr, planter, sopp) er dette tilstede i cellekjernen i form av kromosomer. Et kromosom består av et enkelt, sammenhengende DNA-molekyl, som er knyttet til visse proteiner.
Navnet kromosom er avledet fra gresk og kan grovt sett oversettes som "fargekropp". Dette navnet kommer fra det faktum at forskere lyktes veldig tidlig i cytologihistorien (1888) å fargelegge dem ved hjelp av spesielle grunnleggende fargestoffer og identifisere dem i et lysmikroskop. Imidlertid er de bare veldig synlige på et bestemt punkt i cellesyklusen, mitose (i kimceller, meiose), når kromosomet er spesielt tett (kondensert).

Hvordan bygges kromosomer?
Hvis hele DNA-dobbeltspiralen til en celle, dvs. rundt 3,4 x 109 basepar, ble koblet sammen, ville dette resultere i en lengde på over en meter. Den totale lengden på alle tilsatte kromosomer er bare ca. 115 µm. Denne forskjellen i lengde forklares av den meget kompakte strukturen til kromosomene, der DNA blir viklet eller spiralformet flere ganger på en veldig spesifikk måte.
Histoner, en spesiell form for proteiner, spiller en viktig rolle i dette. Det er totalt 5 forskjellige histoner: H1, H2A, H2B, H3 og H4. To av de siste fire histonene kombineres for å danne en sylindrisk struktur, oktameren, rundt hvilken dobbel helix snor seg rundt to ganger (= super helix). H1 fester seg til denne strukturen for å stabilisere den.
Dette komplekset av DNA, oktamer og H1 kalles et nukleosom. Flere av disse nukleosomene er nå "som en perlestreng" med relativt korte intervaller (10-60 basepar) etter hverandre. Seksjonene mellom kromosomene kalles spacer DNA. De enkelte nukleosomene kommer nå i kontakt igjen via H1, noe som skaper en ytterligere spiral og dermed også en kompresjon.
Den resulterende strengen er i sin tur i løkker som er stabilisert av en ryggrad laget av sure ikke-histonproteiner, også kjent som hertoner. Disse sløyfene er i sin tur til stede i spiraler som er stabilisert av proteiner, noe som resulterer i siste trinn av kompresjon. Imidlertid oppstår denne høye komprimeringsgraden bare i sammenheng med celledeling under mitose.
I denne fasen kan du også se den karakteristiske formen til kromosomene, som består av to kromatider. Stedet hvor disse er koblet til kalles sentromeren. Den deler hvert metafasekromosom i to korte og to lange armer, også kalt p- og q-armer.
Hvis sentromeren ligger omtrent i midten av kromosomet, kalles det et metasentrisk kromosom, hvis det ligger helt i den ene enden av et akrosentrisk kromosom. De i mellom kalles submetasentriske kromosomer. Disse forskjellene, som allerede kan sees under lysmikroskopet, sammen med lengden, tillater en første klassifisering av kromosomene.
Hva er telomerer?
Telomerer er endene på kromosomene med gjentatte sekvenser (TTAGGG). Disse inneholder ikke relevant informasjon, men tjener til å forhindre tap av mer relevante DNA-seksjoner. For hver celledeling går en del av kromosomet tapt gjennom mekanismen for DNA-replikasjon.
Så telomerer er på en måte en buffer som forsinker punktet der cellen mister viktig informasjon ved å dele. Hvis telomerer i en celle er mindre enn 4000 basepar i lengde, startes programmert celledød (apoptose). Dette forhindrer spredning av defekt genetisk materiale i organismen. Noen få celler har telomeraser, dvs. enzymer som er i stand til å forlenge telomerene igjen.
I tillegg til stamcellene, som alle andre celler kommer fra, er dette kimceller og visse celler i immunsystemet. I tillegg finnes også telomeraser i kreftceller, og det er derfor man snakker om udødeliggjøring av en celle i denne sammenhengen.
Les alt om temaet her: Telomerer - Anatomi, funksjon og sykdommer
Hva er kromatin?
Kromatin refererer til hele innholdet i en cellekjerne som kan farges med en base. Derfor inkluderer begrepet i tillegg til DNA også visse proteiner, for eksempel histoner og hertoner (se struktur), så vel som visse RNA-fragmenter (hn og snRNA).
Avhengig av fasen i cellesyklusen eller avhengig av den genetiske aktiviteten, er dette materialet tilgjengelig i forskjellige tettheter. Den tettere formen kalles heterokromatin. For å gjøre det lettere å forstå, kunne man derfor betrakte det som en "lagringsform" og her igjen skille mellom konstituerende og fakultativ heterokromatin.
Konstitutivt heterokromatin er den tetteste formen, som er tilstede i alle faser av cellesyklusen i sitt høyeste kondensnivå. Den utgjør omtrent 6,5% av det menneskelige genomet og ligger hovedsakelig nær sentromerer og ender av kromosomarmene (telomerer) i liten grad, men også andre steder (hovedsakelig kromosomene 1, 9, 16, 19 og Y) . I tillegg er det meste av det konstituerende heterokromatinet lokalisert nær kjernemembranen, dvs. ved kantene av cellekjernen. Plassen i midten er reservert for det aktive kromatinet, eukromatinet.
Fakultativ heterokromatin er litt mindre tett og kan aktiveres og deaktiveres etter behov eller avhengig av utviklingsstadiet. Et godt eksempel på dette er det andre X-kromosomet i kvinnelige karyotyper. Siden ett X-kromosom i utgangspunktet er tilstrekkelig for at cellen skal overleve, slik det til slutt er tilstrekkelig for menn, blir en av de to deaktivert i embryonale faser. det deaktiverte X-kromosomet er kjent som Barr-kroppen.
Bare under celledeling, i sammenheng med mitose, kondenserer den fullstendig, hvorved den når sin høyeste kompresjon i metafasen. Men siden de forskjellige genene leses forskjellig ofte - tross alt kreves ikke hvert protein i samme mengde til enhver tid - det skilles også her mellom aktivt og inaktivt eukromatin.
Les mer om dette under: Kromatin
Haploide kromosomer
Haploid (gresk haploos = enkelt) betyr at alle kromosomer i en celle er tilstede hver for seg, dvs. ikke i par (diploide) som det vanligvis er tilfelle. Dette er den naturlige tilstanden til alle egg- og sædceller, der de to identiske kromatidene ikke ble separert i løpet av den første meiosen, men i stedet skilles alle par av kromosomer først.
Som et resultat, etter den første meiosen, har dattercellene hos mennesker bare 23 i stedet for de vanlige 46 kromosomene, noe som tilsvarer halvparten av det haploide settet med kromosomer. Siden disse dattercellene fremdeles har en identisk kopi av hvert kromosom som består av 2 kromosomer, kreves den andre meiosen, der de to kromatidene er skilt fra hverandre.
Polytene-kromosomer
Et polytene-kromosom er et kromosom som består av et stort antall genetisk identiske kromatider. Siden slike kromosomer er lette å se selv under en lavere forstørrelse, blir de noen ganger referert til som gigantiske kromosomer. Forutsetningen for dette er endoreplikering, der kromosomene i cellekjernen multipliseres flere ganger uten at celledeling forekommer.
Hva er funksjonene til kromosomene?
Kromosomet som organisasjonsenhet for vårt genom tjener primært til å sikre at det doblede genomet fordeles jevnt mellom dattercellene under celledeling. For å gjøre dette er det verdt å se nærmere på mekanismene for celledeling eller cellesyklus:
Cellen tilbringer mesteparten av cellesyklusen i mellomfasen, noe som betyr hele tidsperioden der cellen ikke er i ferd med å dele seg. Dette er igjen delt inn i G1, S og G2 faser.
G1-fasen (G som i gap, dvs. gap) følger umiddelbart celledeling. Her øker cellen i størrelse igjen og utfører generelle metabolske funksjoner.
Herfra kan den også bytte til G0-fasen. Dette betyr at det skifter til et stadium som ikke lenger er i stand til å dele seg, og i normale tilfeller endres også sterkt for å oppfylle en veldig spesifikk funksjon (celledifferensiering). For å oppfylle disse oppgavene leses veldig spesifikke gener mer intenst, andre mindre eller ikke i det hele tatt.
Hvis et segment av DNA ikke er nødvendig over lang tid, er det ofte plassert i de delene av kromosomene som har vært tett pakket i lang tid (se kromatin). På den ene siden har dette som formål å spare plass, men i tillegg til de andre mekanismene for genregulering, er det også en ekstra beskyttelse mot å bli lest ved et uhell. Imidlertid har det også blitt observert at differensierte celler fra G0-fasen under veldig spesifikke forhold kan komme inn i syklusen igjen.
G1-fasen etterfølges av S-fasen, dvs. fasen der nytt DNA syntetiseres (DNA-replikasjon). Her må hele DNA'et være i sin løseste form, dvs. alle kromosomer er helt uviklet (se struktur).
På slutten av syntesefasen er hele genetisk materiale tilstede i duplikat i cellen. Siden kopien fremdeles er festet til det originale kromosomet via sentromeren (se struktur), snakker man ikke om en duplisering av kromosomene.
Hvert kromosom består nå av to kromatider i stedet for ett, slik at det senere kan få den karakteristiske X-formen under mitose (strengt tatt gjelder X-formen bare for metasentriske kromosomer). I den påfølgende G2-fasen foregår den umiddelbare forberedelsen for celledeling. Dette inkluderer også en detaljert kontroll for replikasjonsfeil og strandbrudd, som kan repareres om nødvendig.
Det er i utgangspunktet to typer celledeling: mitose og meiose. Med unntak av kimcellene oppstår alle celler i en organisme gjennom mitose, hvis eneste oppgave er dannelsen av to genetisk identiske datterceller.
Meiose, derimot, har som formål å generere genetisk forskjellige celler:
I et første trinn deles de tilsvarende (homologe) men ikke identiske kromosomene opp. Bare i neste trinn blir kromosomene, som består av to identiske kromatider, separert og igjen distribuert til to datterceller hver, slik at til slutt fire kimceller med ulikt genetisk materiale oppstår fra en forløpercelle.
Formen og strukturen til kromosomene er avgjørende for begge mekanismene: Spesielle "proteintråder", det såkalte spindelapparatet, fester seg til de høyt kondenserte kromosomene og trekker kromosomene i en finregulert prosess fra midtplanet (ekvatorialplan) til motsatte poler av cellen rundt en for å sikre jevn fordeling. Selv små endringer i mikrostrukturen til kromosomene kan få alvorlige konsekvenser her.
I alle pattedyr bestemmer forholdet mellom kjønnskromosomene X og Y også avkomets kjønn. I utgangspunktet avhenger alt av om sædcellene som forener seg med eggcellen har et X- eller et Y-kromosom. Siden begge sædformene alltid produseres i nøyaktig samme grad, er sannsynligheten alltid balansert for begge kjønn. Dette tilfeldige systemet garanterer en jevnere kjønnsfordeling enn hva som for eksempel vil være med miljøfaktorer som temperatur.
Finn ut mer om temaet: Cellekjernedeling
Hvordan videreføres den genetiske sammensetningen via kromosomene?
I dag vet vi at egenskaper arves via gener som lagres i cellene i form av DNA. Disse er i sin tur delt inn i 46 kromosomer, hvor de 25.000-30000 menneskelige genene distribueres.
I tillegg til selve egenskapen, som kalles fenotypen, er det også den genetiske ekvivalenten, som kalles genotypen. Stedet hvor et gen er på et kromosom kalles et lokus. Siden mennesker har dobbelt hvert kromosom, forekommer hvert gen også to ganger. Det eneste unntaket fra dette er X-kromosomgenene hos menn, ettersom Y-kromosomet bare bærer en brøkdel av den genetiske informasjonen som finnes på X-kromosomet.
Ulike gener som er på samme sted kalles alleler. Ofte er det mer enn to forskjellige alleler på ett sted. Man snakker da om polymorfisme. En slik allel kan rett og slett være en ufarlig variant (normal variant), men også patologiske mutasjoner som kan være utløseren for en arvelig sykdom.
Hvis mutasjonen av et enkelt gen er tilstrekkelig til å endre fenotypen, snakker man om monogen eller mendelsk arv. Imidlertid er mange av de arvelige egenskapene arvet gjennom flere interagerende gener og er derfor mye vanskeligere å studere.
Siden mor og far hver overfører et av sine to gener til barnet i en mandelsk arv, er det alltid fire mulige kombinasjoner i neste generasjon, hvor disse også kan være de samme i forhold til en eiendom. Hvis begge allelene til et individ har samme effekt på fenotypen, er individet homozygot i forhold til denne karakteristikken, og karakteristikken blir tilsvarende fullt uttrykt.
Heterozygoter har to forskjellige alleler som kan samhandle med hverandre på forskjellige måter: Hvis en allel er dominerende over en annen, undertrykker den fullstendig uttrykket og den dominerende egenskapen blir synlig i fenotypen. Den undertrykte allelen kalles recessiv.
Når det gjelder en kodominant arv, kan begge alleler uttrykke seg upåvirket av hverandre, mens det i tilfelle en mellomarv er en blanding av begge kjennetegn. Et godt eksempel på dette er blodgruppesystemet AB0, der A og B er dominerende med hverandre, men 0 dominerer over hverandre.
Hva er det normale settet med kromosomer hos mennesker?
Menneskelige celler har 22 kjønnsuavhengige par kromosomer (autosomer) og to kjønnskromosomer (gonosomer), så totalt 46 kromosomer utgjør ett sett kromosomer.
Autosomes kommer vanligvis i par. Kromosomene til et par er like i form og sekvens av gener og blir derfor referert til som homologe. De to X-kromosomene hos kvinner er også homologe, mens menn har et X- og et Y-kromosom. Disse varierer i form og antall gener som er tilstede på en slik måte at man ikke lenger kan snakke om homologi.
Kjønnsceller, dvs. egg- og sædceller, har bare halvparten av kromosomsettet på grunn av meiose, nemlig 22 individuelle autosomer og ett gonosom hver. Siden kimcellene smelter sammen under befruktning og noen ganger bytter hele segmenter (crossover), opprettes en ny kombinasjon av kromosomer (rekombinasjon). Alle kromosomer sammen kalles karyotypen, som med noen få unntak (se kromosomavvik) er identisk hos alle personer av samme kjønn.
Her kan du finne ut alt om emnet: Mitosis - Simply Explained!
Hvorfor er det alltid par kromosomer?
I utgangspunktet kan dette spørsmålet besvares med en setning: Fordi det har vist seg å være gunstig.Tilstedeværelsen av kromosompar og prinsippet om rekombinasjon er avgjørende for arv når det gjelder seksuell reproduksjon. På denne måten kan et helt nytt individ tilfeldigvis dukke opp fra genetisk materiale til to individer.
Dette systemet øker enormt mangfoldet av egenskaper i en art og sørger for at det kan tilpasse seg endrede miljøforhold mye raskere og mer fleksibelt enn det bare ville være mulig ved hjelp av mutasjon og seleksjon.
Det doble settet med kromosomer har også en beskyttende effekt: hvis en mutasjon av et gen vil føre til en funksjonsfeil, er det fortsatt en slags "sikkerhetskopi" i det andre kromosomet. Dette er ikke alltid nok for at organismen skal kompensere for funksjonsfeil, spesielt hvis den muterte allelen er dominerende, men det øker sjansen for det. I tillegg videreføres ikke denne mutasjonen automatisk til alle avkom, som igjen beskytter arten mot altfor radikale mutasjoner.
Hva er en kromosommutasjon?
Genetiske defekter kan oppstå fra ioniserende stråling (f.eks. Røntgenstråler), kjemiske stoffer (f.eks. Benzopyren i sigarettrøyk), visse virus (f.eks. HP-virus) eller, med lav sannsynlighet, kan de også oppstå rent tilfeldig. Det er ofte flere faktorer involvert i utviklingen. I prinsippet kan slike endringer forekomme i alle kroppsvev, men av praktiske årsaker er analysen vanligvis begrenset til lymfocytter (en spesiell type immuncelle), fibroblaster (bindevevsceller) og benmargsceller.
En kromosommutasjon er en stor strukturell endring i individuelle kromosomer. Fravær eller tilsetning av hele kromosomer, derimot, ville være et genom eller ploidemutasjon, mens begrepet genmutasjon refererer til relativt små endringer i et gen. Begrepet kromosomavvik (latinsk aberrare = å avvike) er noe bredere og inkluderer alle endringer som kan oppdages med lysmikroskopet.
Mutasjoner kan ha veldig forskjellige effekter:
- Stille mutasjoner, dvs. mutasjoner der endringen ikke har noen innvirkning på individet eller deres avkom, er ganske atypiske for kromosomavvik og er oftere funnet i området for gen- eller punktmutasjoner.
- Man snakker om en tap av funksjonsmutasjon når mutasjonen resulterer i et feilfoldet og derfor funksjonsfritt protein eller ikke noe protein i det hele tatt.
- Såkalte gain-of-function-mutasjoner endrer typen effekt eller mengden proteiner som produseres på en slik måte at helt nye effekter oppstår. På den ene siden er dette en avgjørende mekanisme for evolusjon og dermed for overlevelse av en art eller fremveksten av nye arter, men på den annen side, som i tilfellet med Philadelphia-kromosomet, kan den også gi et avgjørende bidrag til utvikling av kreftceller.
Den mest kjente av de forskjellige formene for kromosomavvik er sannsynligvis de numeriske avvikene, der individuelle kromosomer bare er tilstede en gang (monosomi) eller til og med tredobbelt (trisomi).
Hvis dette bare gjelder et enkelt kromosom, kalles det aneuploidi, og hele kromosomsettet påvirkes av polyploidi (tri- og tetraploidi). I de fleste tilfeller oppstår denne feilfordelingen i løpet av kimcellens utvikling gjennom ikke-separasjon (ikke-separasjon) av kromosomene under celledeling (meiose). Dette fører til en ujevn fordeling av kromosomene på dattercellene og dermed til numerisk aberrasjon hos barnet.
Monosomier av ikke-kjønnede kromosomer (= autosomer) er uforenlige med livet og forekommer derfor ikke hos levende barn. Med unntak av trisomiene 13, 18 og 21, fører autosomale trisomier nesten alltid til spontane aborter.
I alle fall, i motsetning til avvikene i kjønnskromosomene, som også kan være iøynefallende, er det alltid alvorlige kliniske symptomer og som regel mer eller mindre uttalt ytre abnormiteter (dysmorfier).
En slik maldistribusjon kan også forekomme senere i livet med mitotisk celledeling (alle celler unntatt kimceller). Siden det er uendrede celler i tillegg til de berørte cellene, snakker man om en somatisk mosaikk. Med somatisk (gresk soma = kropp) menes alle celler som ikke er kimceller. Siden bare en liten del av kroppens celler påvirkes, er symptomene vanligvis mye mildere. Derfor forblir mosaiktyper ofte uoppdaget i lang tid.
Her kan du finne ut alt om emnet: Kromosommutasjon
Hva er kromosomavvik?
Den strukturelle kromosomavviket tilsvarer i utgangspunktet definisjonen av kromosommutasjonen (se ovenfor). Hvis mengden genetisk materiale forblir den samme og rett og slett fordeles annerledes, snakker man om en balansert aberrasjon.
Dette gjøres ofte via translokasjon, dvs. overføring av et kromosomsegment til et annet kromosom. Hvis det er en utveksling mellom to kromosomer, snakker man om gjensidig translokasjon. Siden det bare kreves omtrent 2% av genomet for å produsere proteiner, er sannsynligheten veldig lav for at et slikt gen er ved brytpunktet og derved mister sin funksjon eller er svekket i det. Derfor går en slik balansert avvik ofte ubemerket og videreføres i flere generasjoner.
Dette kan imidlertid føre til en maldistribusjon av kromosomer under utvikling av kjønnscellene, noe som kan føre til infertilitet, spontane spontanaborter eller avkom med en ubalansert aberrasjon.
En ubalansert aberrasjon kan også forekomme spontant, dvs. uten familiehistorie. Sannsynligheten for at et barn blir født levende med ubalansert aberrasjon, avhenger sterkt av kromosomene som påvirkes, og varierer mellom 0 og 60%. Dette fører til tap (= sletting) eller duplisering (= duplisering) av et kromosomsegment. I denne sammenhengen snakker man også om delvis mono- og trisomier.
I noen tilfeller forekommer disse sammen i to forskjellige regioner, hvor delvis monosomi vanligvis er mer avgjørende for forekomsten av kliniske symptomer. Dette er fremtredende eksempler på en sletting Cat Scream Syndrome og Wolf-Hirschhorn Syndrome.
Man snakker om en mikrodeletjon når endringen ikke lenger kan bestemmes med lysmikroskopet, dvs. når det dreier seg om tap av ett eller få gener. Dette fenomenet anses å være årsaken til Prader-Willi syndrom og Angelman syndrom og er nært knyttet til utviklingen av retionoblastom.
Robertson-translokasjonen er et spesielt tilfelle:
To akrosentriske kromosomer (13, 14, 15, 21, 22) forenes ved sentromeren og danner et enkelt kromosom etter at kortarmene har gått tapt (se struktur). Selv om dette resulterer i redusert antall kromosomer, blir dette referert til som en balansert aberrasjon, ettersom tapet av korte armer i disse kromosomene lett kan kompenseres for. Også her er effekten ofte bare merkbar i påfølgende generasjoner, da det er svært stor sannsynlighet for spontanabort eller levende barn med trisomi.
Hvis det er to brudd i et kromosom, kan det skje at mellomsegmentet roteres 180 ° og inkorporeres i kromosomet. Denne prosessen, kjent som inversjon, er bare ubalansert hvis bruddpunktet ligger i et aktivt gen (2% av det totale genetiske materialet). Avhengig av om sentromeren er innenfor eller utenfor det inverterte segmentet, er det en peri- eller parasentrisk inversjon. Disse endringene kan også bidra til ujevn fordeling av genetisk materiale på kimcellene.
I parasentrisk inversjon, der sentromeren ikke er i det inverterte segmentet, kan også kimceller med to eller ingen sentromere vises. Som et resultat går det tilsvarende kromosomet tapt under de aller første celledelingene, noe som nesten helt sikkert fører til spontanabort.
Innsetting er installasjon av et kromosomfragment andre steder. Også her blir avkommet først og fremst påvirket på en lignende måte. Et ringkromosom kan forekomme spesielt etter sletting av endestykker. Type og størrelse på sekvensene er avgjørende for alvorlighetsgraden av symptomene. I tillegg kan dette føre til feil fordeling og dermed igjen til mosaiktyper i kroppens celler.
Hvis metafasekromosomet skilles feil under celledeling, kan det oppstå isokromosomer. Dette er to nøyaktig de samme kromosomene som bare består av lange eller bare korte armer. I tilfelle av X-kromosomet kan dette manifestere seg som et Ulrich-Turners syndrom (monosomi X).
Les mer informasjon om dette emnet: Kromosomavvik
Trisomi 21
Trisomi 21, bedre kjent som Downs syndrom, er trolig den vanligste numeriske kromosomavviket blant levendefødte, og menn blir litt oftere rammet (1.3: 1).
Sannsynligheten for at trisomi 21 skal oppstå avhenger av forskjellige demografiske faktorer, som gjennomsnittsalderen ved fødselen til mødrene, og varierer litt fra region til region.
95% av trisomien 21 oppstår som et resultat av en delingsfeil i sammenheng med meiose (kimcelledeling), nemlig ikke-disjunksjon, dvs. manglende separasjon av søsterkromatidene.
Disse er kjent som frie trisomier og oppstår 90% i mors, 5% i fader og ytterligere 5% i det embryonale genomet.
Ytterligere 3% resultat av ubalanserte translokasjoner enten på kromosom 14 eller som 21; 21 translokasjon, og skaper et normalt og dobbelt kromosom 21. De resterende 2% er mosaiktyper der trisomien ikke oppsto i kimceller og derfor ikke påvirker alle kroppens celler. Mosaiktyper er ofte så milde at de kan forbli helt uoppdaget i lang tid.
Uansett bør det utføres en kromosomundersøkelse for å skille symptomatisk identisk fri trisomi fra den muligens arvelige translokasjonstrisomi. En familiehistorie fra tidligere generasjoner kan deretter følge.
Er du interessert i dette emnet? Les neste artikkel om dette: Trisomi 21
Trisomi 13
Trisomy 13 eller Patau syndrom har en frekvens på 1: 5000 og er mye sjeldnere enn Downs syndrom. Årsakene (gratis trisomier, translokasjoner og mosaikktyper) og deres prosentvise fordeling er stort sett identiske.
I teorien kan nesten alle tilfeller diagnostiseres prenatalt ved hjelp av ultralyd eller PAPP-A-testen. Siden PAPP-A-testen ikke nødvendigvis er en del av rutinemessige undersøkelser, blir rundt 80% av tilfellene i Sentral-Europa diagnostisert før fødselen.
En vekstrest, en bilateral leppe og gane i spalten og uvanlig små øyne (mikroftalmi) kan allerede sees i ultralyden. I tillegg er misdannelser i forhjernen og ansiktet av forskjellige alvorlighetsgrader vanligvis til stede (holoprosensefali).
Mens hjernehalvkulene i lobarformen er nesten helt atskilt, og det blir opprettet laterale ventrikler, er det i semi-lobarformen ofte bare den bakre delen av hjernen atskilt og de laterale ventriklene mangler. I den mest alvorlige formen, alobarformen, er det ingen separasjon av hjernehalvkulene.
Spedbarn med halv- eller alobarform dør vanligvis umiddelbart etter fødselen. Etter en måned er dødeligheten rundt 50% av levendefødte. Inntil 5 år øker dødeligheten fra trisomi 13 til 90%. På grunn av misdannelser i hjernen, forblir syke mennesker i de fleste tilfeller sengeliggende hele livet og kan ikke snakke, og det er derfor de er avhengige av full pleie. I tillegg kan det også være vidtrekkende fysiske manifestasjoner av Trismoie 13.
Les mer om emnet på: Trisomi 13 hos det ufødte barnet
Trisomi 16
I utgangspunktet er trisomi 16 den vanligste trisomien (rundt 32% av alle trisomier), men levende barn med trisomi 16 er svært sjeldne. Generelt forekommer levende fødsler bare i delvis trisomier eller mosaiktyper. Blant trisomiene er det imidlertid hyppigst ansvarlig for dødfødsler: 32 av 100 spontanaborter på grunn av kromosomavvik kan spores tilbake til denne formen for trisomi.
Derfor er hovedsakelig prenatal, dvs. prenatal, identifiserbare egenskaper blitt dokumentert. Bemerkelsesverdig her er forskjellige hjertefeil, redusert vekst, en enkelt navlearterie (ellers dobbelt) og økt nakketransparens, noe som forklares med væskeansamling på grunn av det ennå ikke fullt utviklede lymfesystemet og den økte elastisiteten i huden i dette området. I tillegg vil den fysiologiske navlestrengen, dvs. den midlertidige forskyvningen av en stor del av tarmen gjennom navlen til utsiden, ofte ikke trekke seg ordentlig tilbake, noe som er kjent som en omphalocele eller navlestrengsbrudd.
En fleksjonskontraktur med kryssede fingre kan også ofte oppdages ved ultralyd. Hos noen få fødsler er generalisert muskelhypotensjon, dvs. generell muskelsvakhet, merkbar. Dette fører til svakhet ved drikking og kan sikre at spedbarnet må mates kunstig. Firefingersfuren som er så karakteristisk for trisomier forekommer også ofte. Også her er hyppigheten av forekomsten av trisomien direkte relatert til morens alder.
Trisomi 18
Edwards syndrom, dvs. trisomi 18, forekommer med en frekvens på 1: 3000. Med fosterdiagnostikk er det det samme som med Patau syndrom: Også her vil de samme undersøkelsene tillate at alle pasientene blir funnet helt før fødselen. Årsakene og fordelingen av dem kan sammenlignes med andre trisomier (se trisomi 21).
I tillegg forekommer partielle trisomier i trisomi 18, som i likhet med mosaiktypene fører til mye mildere kliniske kurs. De tilknyttede dysmorfismene er også ekstremt karakteristiske for Edwards syndrom: Ved fødselen har pasienter en sterkt redusert kroppsvekt på 2 kg (normal: 2,8-4,2 kg), en tilbaketrekkende bred panne, en generelt underutviklet nedre halvdel av ansiktet med en liten munn åpning, smale øyelokkespalter og roterte bakover, formendrede ører (faunens øre). Baksiden av hodet, som er uvanlig sterkt utviklet for en nyfødt, er også merkbar. Ribbeina er uvanlig smale og skjøre. Nyfødte har også en permanent spenning (tone) i hele muskulaturen, som imidlertid trekker seg tilbake hos de overlevende etter de første ukene.
Et annet karakteristisk trekk er kryssing av andre og femte fingre over 3. og 4. med totalt antall fingre som er hamret inn, mens føttene er uvanlig lange (forløpne), har en spesielt uttalt hæl, stuntede tånegler og en tilbaketrukket stortå .
Alvorlige organdeformasjoner er vanlige og forekommer vanligvis i kombinasjon: hjerte- og nyredefekter, malfolding (malrotasjon) i tarmen, vedheft i bukhinnen (mesenterium kommune), en okklusjon av spiserøret (esophageal atresia) og mange flere.
På grunn av disse misdannelsene er dødeligheten rundt 50% i løpet av de første 4 dagene, bare rundt 5-10% lever for å være over ett år gammel. Overlevelse til voksen alder er det absolutte unntaket. Uansett er en intellektuell funksjonshemming veldig uttalt og kan ikke snakke, er sengeliggende og inkontinent, så helt avhengig av hjelp utenfor.
For mer detaljert informasjon om trisomi 18, vennligst les også vår detaljerte artikkel om emnet:
- Trisomi 18 (Edwards syndrom)
- Trisomi 18 hos det ufødte barnet
Trisomi X
Trisomi X er den mest iøynefallende formen for numerisk kromosomavvik, utseendet til de berørte, som logisk sett er alle kvinner, skiller seg ikke sterkt fra andre kvinner. Noen skiller seg ut fordi de er spesielt høye og har noe “fyldige” ansiktsegenskaper. Mental utvikling kan også i stor grad være normal, alt fra borderline normal til mild mental funksjonshemming.
Imidlertid er dette intelligensunderskuddet noe mer alvorlig enn med de andre trisomiene i kjønnskromosomene (XXY og XYY). Med en frekvens på 1: 1000 er det faktisk ikke så sjelden, men siden trisomi vanligvis ikke er forbundet med klinisk signifikante symptomer, vil sannsynligvis flertallet av kvinnene med sykdommen aldri bli diagnostisert hele livet.
Bærere blir for det meste oppdaget ved en tilfeldighet under familiekontroll eller under prenatal diagnostikk.Fertiliteten kan reduseres noe, og frekvensen av kjønns kromosomavvik i neste generasjon kan økes noe, slik at genetisk rådgivning anbefales hvis du ønsker å få barn.
Som med de andre trisomiene, utvikler trisomi X oftest seg som en fri trisomi, dvs. på grunn av mangel på splittelse (søvnkromatidene). Også her oppstår det vanligvis under modningen av mors eggceller, selv om sannsynligheten øker med alderen.
Fragile X Syndrome
Fragile X-syndrom eller Martin Bells syndrom foretrekkes hos menn, ettersom de bare har ett X-kromosom og derfor er mer påvirket av endringen.
Det forekommer med en frekvens på 1: 1250 blant levende mannlige fødsler på ett år, noe som gjør den til den vanligste formen for uspesifikk mental retardasjon, dvs. alle mentale funksjonshemninger som ikke kan beskrives av et spesielt syndrom med typiske tegn.
Fragile X syndrom kan vanligvis forekomme hos jenter i en noe svakere form, noe som skyldes utilsiktet inaktivering av en av X-kromosomene. Jo høyere andel av det avslåtte sunne X-kromosomet, desto sterkere er symptomene.
Mesteparten av tiden er imidlertid kvinner bærere av premutasjonen, som foreløpig ikke gir noen kliniske symptomer, men som øker sannsynligheten for full mutasjon hos sønnene. I svært sjeldne tilfeller kan menn også være bærere av premutasjonen, som de da bare kan videreføre til døtre som imidlertid vanligvis er klinisk sunne (Sherman paradox).
Syndromet utløses av et ekstremt økt antall CGG-tripletter (en viss basesekvens) i FMR-genet (skjørt sted-mental retardasjon); i stedet for 10-50 eksemplarer, premutasjonen 50-200, når den er fullt utviklet 200- 2000 eksemplarer.
Under lysmikroskopet ser dette ut som et brudd i den lange armen, og det er det som ga syndromet navnet. Dette fører til deaktivering av det berørte genet, som igjen forårsaker symptomene.
Berørte mennesker viser en bremset utvikling av tale og bevegelse og kan vise atferdsproblemer som kan gå i retning av hyperaktivitet, men også autisme. Rent ytre abnormiteter (tegn på dysmorfisme) er et langt ansikt med en fremtredende hake og utstående ører. Med puberteten blir testiklene ofte sterkt forstørret (makroorchidia) og ansiktsegenskapene blir grovere. Det er en liten opphopning av psykologiske abnormiteter og en særlig tidlig overgangsalder blant kvinnelige bærere av premutasjonen.
Hva er en kromosomanalyse?
Kromosomanalyse er en prosess i cytogenetikk som numeriske eller strukturelle kromosomavvik kan oppdages.
En slik analyse vil for eksempel bli brukt hvis det umiddelbart mistenkes et kromosomalt syndrom, dvs. i tilfelle misdannelser (dysmorfier) eller intellektuelle funksjonshemninger (retardasjon), men også i tilfelle infertilitet, regelmessige aborter (aborter) og også med visse kreftformer (f.eks. lymfomer eller leukemi).
Dette krever vanligvis lymfocytter, en spesiell type immuncelle som oppnås fra pasientens blod. Siden bare en relativt liten mengde kan oppnås på denne måten, stimuleres cellene til å dele seg med fytohemagglutinin, og lymfocyttene kan deretter dyrkes i laboratoriet.
I noen tilfeller tas prøver (biopsier) i stedet fra huden eller ryggmargen, med en lignende prosedyre. Målet er å skaffe så mye DNA-materiale som mulig som for tiden er midt i celledeling. I metafasen er alle kromosomer arrangert i ett nivå omtrent midt i cellen, for å bli trukket til motsatte sider (poler) av cellen i neste trinn, anafasen.
På dette tidspunktet er kromosomene spesielt tett pakket (høyt kondensert). Spindelgiften colchicine tilsettes, som fungerer presist i denne fasen av cellesyklusen, slik at metafasekromosomene akkumuleres. De blir deretter isolert og farget ved hjelp av spesielle fargemetoder.
Den vanligste er GTG-banding, der kromosomene blir behandlet med trypsin, et fordøyelsesenzym og pigmentet Giemsa. De spesielt tettpakkete områdene og de som er rike på adenin og tymin er vist mørke.
De resulterende G-båndene er karakteristiske for hvert kromosom og anses forenklet å være regionene med færre gener. Et bilde av kromosomene som er farget på denne måten, er tatt i tusen ganger forstørrelse, og et karyogram opprettes ved hjelp av et dataprogram. I tillegg til båndmønsteret, brukes kromosomstørrelsen og sentromerens plassering for å hjelpe til med å ordne kromosomene deretter. Men det er også andre metoder for banding som kan ha svært forskjellige fordeler.
Anbefalinger fra redaksjonen
For mer generell informasjon, se følgende artikler:
- Cellekjernedeling
- Funksjoner i cellekjernen
- Trisomi 21
- Genetiske sykdommer


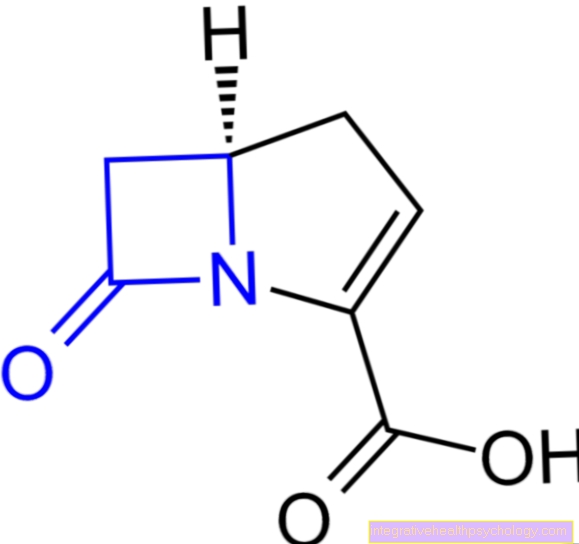














.jpg)